Морфо-функциональная характеристика и классификация хромосом
Ядерная оболочка
Состоит из двух мембран - наружной и внутренней. Пространство между ними называется перинуклеарным. Внешняя мембрана может переходить в мембраны ЭПС. Это позволяет постоянно обмениваться содержимым перинуклеарного пространства и каналов ЭПС. Кроме того, наружная мембрана способна формировать пузырьки, которые встраиваются в КГ. Возможен и обратный процесс – пузырьки, сформированные КГ, включаются в ядерную мембрану, изливая содержимое в перинуклеарное пространство.
Ядерная мембрана пронизана порами, через которые из ядра в цитоплазму транспортируются молекула иРНК, сложные образования из рРНК и белков (предшественники рибосом) и некоторые вещества. Пора представляет собой сложный комплекс, состоящий из нескольких белковых глобул.
Изнутри поверхность внутренней ядерной мембраны подстилает белковая сеть – ядерная ламина, которая участвует в организации хроматина и порового комплекса. При делении клетки ламина участвует в разрушении ядерной оболочки.
2.8.2. Нуклеоплазма (кариолимфа, ядерный сок) - внутренняя среда клетки
В виде бесструктурной массы окружает хромосомы и ядрышко. Вязкость кариоплазмы примерно такая же, как и гиалоплазмы, а кислотность выше. Она состоит из воды (основная часть) и растворённых в ней органических и неорганических соединений. Предполагается, что в кариоплазме происходят специфические биохимические процессы.
В зависимости от периода клеточного цикла хромосомы могут находиться в ядре в двух состояниях – конденсированном, частично конденсированном и полностью конденсированном.
Раньше для обозначения упаковки хромосом употребляли термин – спирализация, деспирализация. В настоящее время используют более точный термин конденсация, деконденсация. Этот термин более ёмкий и включает процесс спирализации хромосомы, её укладку и укорочение.
Во время интерфазы экспрессия (функция, работа) генов максимальна и хромосомы имеют вид тонких нитей. Те участки нити, в которых происходит синтез РНК – деконденсированы, а те участки, где синтез не происходит, – наоборот, конденсированы (рис. 19).
Во время деления, когда ДНК в хромосомах практически не функционирует, хромосомы представляют собой плотные тельца, похожие на «Х» или «У». Это связано с сильной конденсацией ДНК в хромосомах.
Особо необходимо уяснить, что наследственный материал по-разному представлен в клетках, находящихся в интерфазе и в момент деления. В интерфазе в клетке отчётливо просматривается ядро, наследственный материал, в котором представлен хроматином. Хроматин, в свою очередь, состоит из частично конденсированных нитей хромосом. Если же рассматривать клетку во время деления, когда ядра уже нет, то весь наследственный материал концентрируется в хромосомах, которые максимально конденсированы (рис. 20).
Совокупность всех нитей хромосом, состоящих из ДНК и различных белков, в ядрах эукариотических клеток носит название хроматин (см. рис. 19. В). Хроматин в свою очередь делится на эухроматин и гетерохроматин. Первый слабо окрашивается красителями, т.к. содержит тонкие неконденсированные нити хромосом. Гетерохроматин, напротив, – содержит конденсированную, а следовательно, хорошо прокрашиваемую нить хромосомы. Неконденсированные участки хроматина содержат ДНК, в которой функционируют гены (т.е. происходит синтез РНК).
Клетка в интерфазе Клетка во время деления
Ядро Хромосомы
А Б
Рис. 20. Два состояния наследственного материала в клетках в клеточном цикле: А – в интерфазе наследственный материал расположен в хромосомах, которые частично деконденсированы и расположены в ядре; Б – при делении клетки наследственный материал выходит из ядра, хромосомы располагаются в цитоплазме.
Необходимо помнить, что если ген функционирует, то ДНК в этом участке деконденсирована. И наоборот, конденсация ДНК гена свидетельствует о блокаде активности гена. Феномен конденсации и деконденсации участков ДНК достаточно часто можно обнаружить, когда в клетке регулируется активность (включения или выключения) генов.
Субмолекулярное строение хроматина (в дальнейшем мы будем их называть интерфазные хромосомы) и хромосом делящейся клетки (в дальнейшем мы будем их называть метафазные хромосомы) до настоящего времени полностью не выяснено. Однако ясно, что при различных состояниях клетки (интерфаза и деление) организация наследственного материала различна. В основе интерфазных (ИХ) и метафазных хромосом (МХ) лежит нуклеосома. Нуклеосома состоит из центральной белковой части, вокруг которой обёрнута нить ДНК. Центральную часть образуют восемь молекул белка-гистона – Н2А, Н2В, Н3, Н4 (каждый гистон представлен двумя молекулами). В связи с этим сердцевина нуклеосомы носит название тетрамер, октамериликор. Молекула ДНК в форме спирали обвивает кор 1,75 раз и переходит на соседний кор, обвивает его и переходит на следующую. Таким образом создаётся своеобразная фигура, напоминающая нитку (ДНК) с нанизанными на ней бусами (нуклеосомами).
Между нуклеосомами лежит ДНК, которая называется линкерной. С ней может связываться ещё один гистон – Н1. Если он связывается с линкерным участком, то ДНК изгибается и сворачивается в спираль (рис. 21. Б). Гистон Н1 принимает участие в сложном процессе конденсации ДНК, при котором нитка бус сворачивается в спираль толщиной 30 нм. Эта спираль носит название соленоид. Нити хромосом интерфазных клеток состоят из нитей бус и соленоидов. В метафазных хромосомах соленоид сворачивается в суперспираль, которая соединяется с сетчатой структурой (из белков), формируя петли, которые укладываются уже в виде хромосомы. Такая упаковка приводит к почти 5000-кратному уплотнению ДНК в метафазной хромосоме. На рисунке 23 представлена схема последовательной укладки хроматина. Понятно, что процесс спирализации ДНК в ИХ и МХ намного сложнее, но сказанное даёт возможность уяснить наиболее общие принципы упаковки хромосом.
Необходимо отметить, что, каждая хромосома в метафазе состоит из двух хроматид, удерживаемых с помощью центромеры (первичной перетяжки). В основе каждой из этих хроматид лежат упакованные порознь дочерние молекулы ДНК. После процесса компактизации они становятся хорошо различимыми в световой микроскоп хроматидами одной хромосомы. В конце митоза они расходятся по дочерним клеткам. С момента отделения хроматид одной хромосомы друг от друга, их уже называют хромосомами, то есть хромосома содержит либо две хроматиды, перед делением, либо – одну (но она называется уже хромосомой) после деления.
Некоторые хромосомы, кроме первичной перетяжки, имеют вторичную. Её ещё называют ядрышковый организатор. Это тонкая нить хромосомы, на конце которой помещается спутник. Вторичная перетяжка, как и основная хромосома, состоит из ДНК, на которой располагаются гены, ответственные за синтез рибосомальных РНК. На концах хромосомы располагается участок, называемый теломерой. Он как бы «запечатывает» хромосому. Если теломера случайно отрывается, образуется «липкий» конец, который может соединиться с таким же концом другой хромосомы.
располагается посередине, хромосома имеет равные по величине плечи. В субметацентрических хромосомах центромера немного сдвинута к одному концу. Плечи хромосомы не одинаковы по длине – одно длиннее другого. В акроцентрических хромосомах центромера располагается почти на конце хромосомы и короткие плечи трудно различимы. Количество хромосом постоянно для каждого вида. Так, кариотип человека содержит 46 хромосом. У дрозофилы их 8, а в клетке пшеницы – 14.
Совокупность всех метафазных хромосом клетки, их форма и морфология называется кариотипом. По форме различают три типа хромосом – метацентрические, субметацентрические и акроцентрические (рис. 23). В метацентрических хромосомах центромера
Это плотное, хорошо прокрашиваемое тельце, расположенное внутри ядра. В нем обнаружены ДНК, РНК и белки. Основу ядрышка составляют ядрышковые организаторы – участки ДНК, несущие множественные копии генов рРНК. На ДНК ядрышковых организаторов происходит синтез рибосомальных РНК. К ним присоединяются белки и формируется сложное образование - рибонуклеопротеидные (РНП) частицы. Это предшественники (или полуфабрикаты) малой и большой субъединиц рибосом. Процесс образования РНП в основном происходит в периферической части ядрышек. Предшественники ри-
Спутник
Вторичная перетяжка
Центромера
А Б В Г
Рис. 23. Строение хромосом человека в метафазе:
А – метацентрическая хромосома, Б – субметацентрическая, В – акроцентрическая, Г – хромосома с вторичной перетяжкой.
босом выходят из ядрышка и через ядерные поры попадают в цитоплазму, где происходит окончательное формирование субъединиц рибосом (рис. 24).
Размер ядрышка отражает степень его функциональной активности, которая широко варьирует в различных клетках и может изменяться в индивидуальной клетке. Чем интенсивнее происходит процесс формирования рибосом в цитоплазме, тем активнее осуществляется синтез специфических белков на рибосомах. В этом отношении примечательно действие стероидных гормонов (СГ) на клетки-мишени. СГ попадают в ядро и активируют синтез рРНК. В результате возрастает количество РНП и, как следствие, увеличивается число рибосом в цитоплазме. Это приводит к значительному возрастанию уровня синтеза специальных белков, которые через ряд биохимических и физиологических реакций обеспечивают определённый фармакологический эффект (например, разрастается железистый эпителий в матке).
В зависимости от фазы клеточного цикла внешний вид ядрышка заметно меняется. С началом митоза ядрышко уменьшается, а затем и вовсе исчезает. В конце митоза, когда возобновляется синтез рРНК, миниатюрные ядрышки вновь возникают на участках хромосом, содержащих гены рРНК.

 Клетка в интерфазе Клетка во время деления
Клетка в интерфазе Клетка во время деления


 Ядро Хромосомы
Ядро Хромосомы
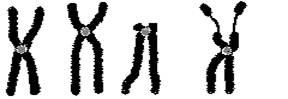 Спутник
Спутник